- Information
- Symbol: MER3,RCK
- MSU: LOC_Os02g40450
- RAPdb: Os02g0617500
- PSP score
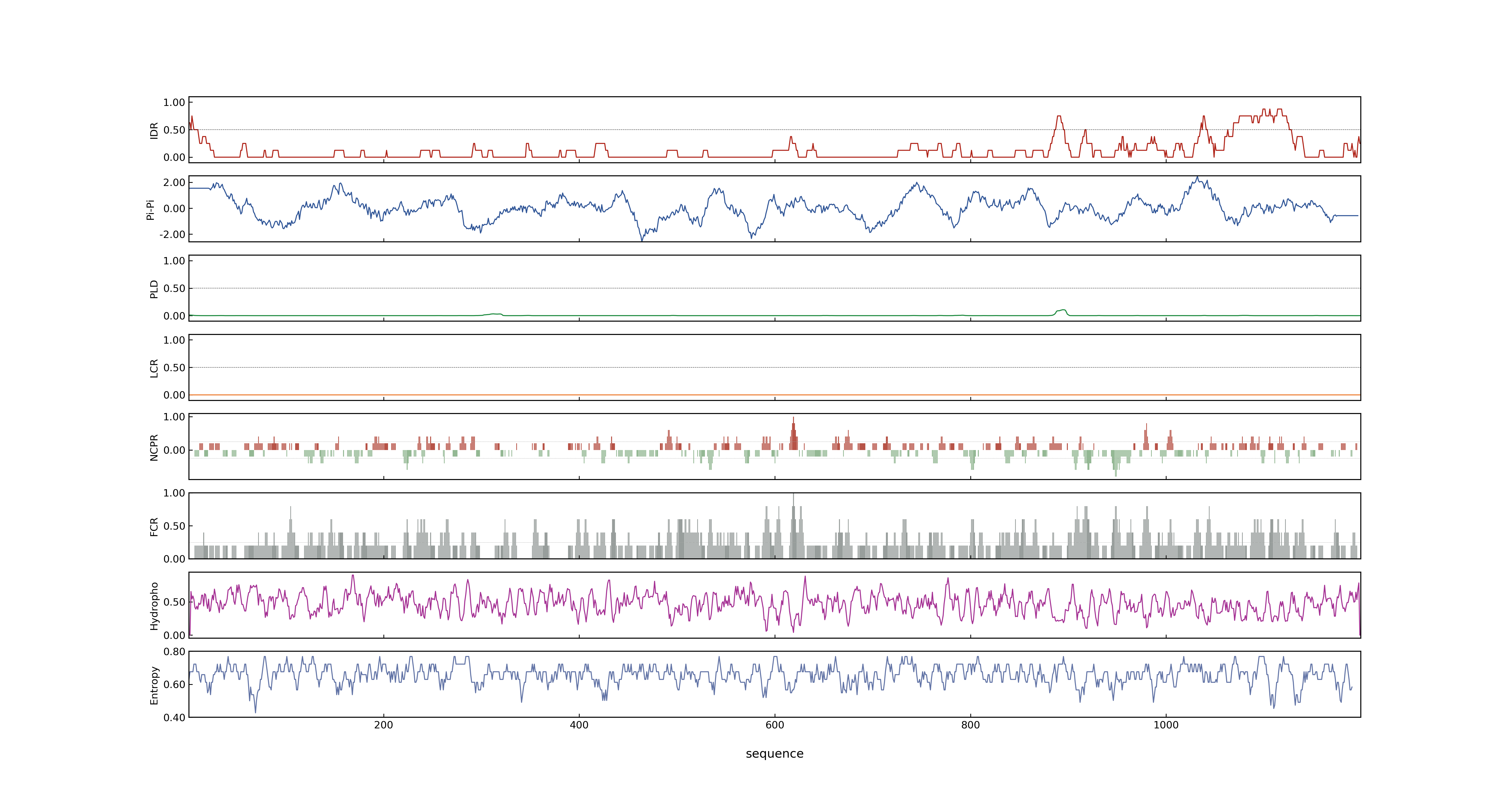
- LOC_Os02g40450.1: 0.0848
- PLAAC score
- LOC_Os02g40450.1: 0
- pLDDT score
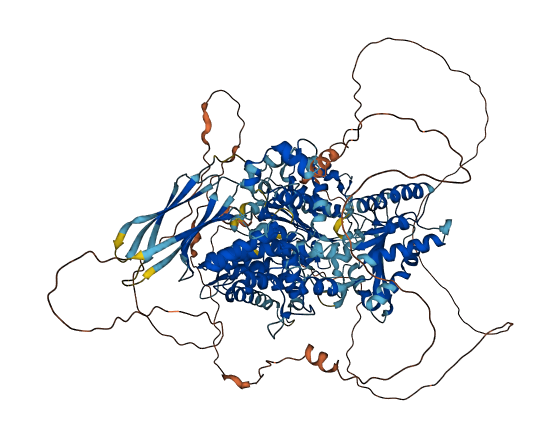
- 72.15
- Protein Structure from AlphaFold and UniProt
- MolPhase score
- LOC_Os02g40450.1: 0.99861681
- MolPhase Result
- Publication
- Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, 2009, Cell Res.
- OsAM1 is required for leptotene-zygotene transition in rice, 2011, Cell Res.
- MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice, 2009, J Cell Sci.
- ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, 2012, J Cell Sci.
- OsSGO1 maintains synaptonemal complex stabilization in addition to protecting centromeric cohesion during rice meiosis, 2011, Plant J.
- The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, 2012, Plant J.
- The role of rice HEI10 in the formation of meiotic crossovers, 2012, PLoS Genet.
- The role of OsMSH5 in crossover formation during rice meiosis, 2013, Mol Plant.
- The central element protein ZEP1 of the synaptonemal complex regulates the number of crossovers during meiosis in rice, 2010, Plant Cell.
- OsREC8 is essential for chromatid cohesion and metaphase I monopolar orientation in rice meiosis, 2011, Plant Physiol.
- OsSPO11-1 is essential for both homologous chromosome pairing and crossover formation in rice, 2010, Chromosoma.
- Genbank accession number
- Key message
- Quantitative real-time RT-PCR analysis indicated that rice SDS and RCK are expressed preferentially in young flowers, and transgenic RNAi rice lines with reduced expression of these genes exhibited normal vegetative development, but showed significantly reduced fertility with partially sterile flowers and defective pollens
- The Arabidopsis SDS (SOLO DANCERS) and RCK (ROCK-N-ROLLERS) genes are important for male meiosis, but it is still unknown whether they represent conserved functions in plants
- Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice
- Finally, we found that the centromeric localization of OsSGO1 depends on OsAM1, not other meiotic proteins such as OsREC8, PAIR2, OsMER3, or ZEP1
- The null mutation of MER3 results in complete sterility without any vegetative defects
- Because rice and Arabidopsis, respectively, are members of monocots and eudicots, two largest groups of flowering plants, our results suggest that the functions of SDS and RCK are likely conserved in flowering plants
- MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice
- Further cytological examination of male meiocytes revealed that the SDS deficiency led to defects in homolog interaction and bivalent formation in meiotic prophase I, and RCK deficiency resulted in defective meiotic crossover formation
- OsMSH5 can be loaded onto meiotic chromosomes in Oszip4, Osmer3, and hei10
- In the absence of OsAM1, many other critical meiotic components, including PAIR2, ZEP1 and OsMER3, could not be correctly installed onto chromosomes
- Several key meiotic proteins, including ZEP1 and OsMER3, were not loaded normally onto chromosomes in Oscom1 mutants, whereas the localization of OsREC8, PAIR2 and PAIR3 seemed to be normal
- Moreover, OsCOM1 was loaded normally onto meiotic chromosomes in Osrec8, zep1 and Osmer3 mutants, but could not be properly loaded in Osam1, pair2 and OsSPO11-1(RNAi) plants
- Cytological analyses show that chiasma frequency is reduced dramatically in mer3 mutants and the remaining chiasmata distribute randomly among different pollen mother cells, implying possible coexistence of two kinds of crossover in rice
- In contrast, in pair2, Osmer3 and zep1 mutants, OsAM1 could be loaded normally, suggesting that OsAM1 plays a fundamental role in building the proper chromosome structure at the beginning of meiosis
- Connection
- MER3~RCK, OsSPO11-1~OsTOP6A1, OsSPO11-1 is essential for both homologous chromosome pairing and crossover formation in rice, The crossover protein, MER3, isn’t efficiently assembled onto chromosomes and the lack of bivalent suggests that crossovers are also affected in the absence of OsSPO11-1
- MER3~RCK, SDS~OsSDS, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, The Arabidopsis SDS (SOLO DANCERS) and RCK (ROCK-N-ROLLERS) genes are important for male meiosis, but it is still unknown whether they represent conserved functions in plants
- MER3~RCK, SDS~OsSDS, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, We have performed phylogenetic analyses of SDS and RCK and their respective homologs, and identified their putative orthologs in poplar and rice
- MER3~RCK, SDS~OsSDS, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, Quantitative real-time RT-PCR analysis indicated that rice SDS and RCK are expressed preferentially in young flowers, and transgenic RNAi rice lines with reduced expression of these genes exhibited normal vegetative development, but showed significantly reduced fertility with partially sterile flowers and defective pollens
- MER3~RCK, SDS~OsSDS, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, Further cytological examination of male meiocytes revealed that the SDS deficiency led to defects in homolog interaction and bivalent formation in meiotic prophase I, and RCK deficiency resulted in defective meiotic crossover formation
- MER3~RCK, SDS~OsSDS, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, These results indicate that rice SDS and RCK genes have similar functions to their Arabidopsis orthologs
- MER3~RCK, SDS~OsSDS, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, Because rice and Arabidopsis, respectively, are members of monocots and eudicots, two largest groups of flowering plants, our results suggest that the functions of SDS and RCK are likely conserved in flowering plants
- MER3~RCK, SDS~OsSDS, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice, Functional conservation of the meiotic genes SDS and RCK in male meiosis in the monocot rice
- MER3~RCK, PAIR2, OsAM1 is required for leptotene-zygotene transition in rice, In the absence of OsAM1, many other critical meiotic components, including PAIR2, ZEP1 and OsMER3, could not be correctly installed onto chromosomes
- MER3~RCK, PAIR2, OsAM1 is required for leptotene-zygotene transition in rice, In contrast, in pair2, Osmer3 and zep1 mutants, OsAM1 could be loaded normally, suggesting that OsAM1 plays a fundamental role in building the proper chromosome structure at the beginning of meiosis
- MER3~RCK, ZEP1, OsAM1 is required for leptotene-zygotene transition in rice, In the absence of OsAM1, many other critical meiotic components, including PAIR2, ZEP1 and OsMER3, could not be correctly installed onto chromosomes
- MER3~RCK, ZEP1, OsAM1 is required for leptotene-zygotene transition in rice, In contrast, in pair2, Osmer3 and zep1 mutants, OsAM1 could be loaded normally, suggesting that OsAM1 plays a fundamental role in building the proper chromosome structure at the beginning of meiosis
- MER3~RCK, OsAM1, OsAM1 is required for leptotene-zygotene transition in rice, In the absence of OsAM1, many other critical meiotic components, including PAIR2, ZEP1 and OsMER3, could not be correctly installed onto chromosomes
- MER3~RCK, OsAM1, OsAM1 is required for leptotene-zygotene transition in rice, In contrast, in pair2, Osmer3 and zep1 mutants, OsAM1 could be loaded normally, suggesting that OsAM1 plays a fundamental role in building the proper chromosome structure at the beginning of meiosis
- MER3~RCK, PAIR2, MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice, In addition, MER3 does not colocalize with PAIR2 at the beginning of prophase I, but locates on one end of PAIR2 fragments at later stages, whereas MER3 foci merely locate on one end of REC8 fragments when signals start to be seen in early prophase I
- MER3~RCK, PAIR2, MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice, The normal loading of PAIR2 and REC8 in mer3 implies that their loading is independent of MER3
- MER3~RCK, PAIR2, MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice, On the contrary, the absence of MER3 signal in pair2 mutants indicates that PAIR2 is essential for the loading and further function of MER3
- MER3~RCK, OsRad21-4~OsREC8, MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice, In addition, MER3 does not colocalize with PAIR2 at the beginning of prophase I, but locates on one end of PAIR2 fragments at later stages, whereas MER3 foci merely locate on one end of REC8 fragments when signals start to be seen in early prophase I
- MER3~RCK, OsRad21-4~OsREC8, MER3 is required for normal meiotic crossover formation, but not for presynaptic alignment in rice, The normal loading of PAIR2 and REC8 in mer3 implies that their loading is independent of MER3
- MER3~RCK, ZEP1, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, Here, we identify the rice ZIP4 homolog, a member of the ZMM gene group, and explore its relationship with two other characterized ZMM genes, MER3 and ZEP1
- MER3~RCK, ZEP1, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, Although zep1 alone gives an increased chiasma number, both zip4 zep1 and mer3 zep1 show a much lower chiasma number than the zip4 or mer3 single mutants
- MER3~RCK, ZEP1, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, These results imply that the normal functions of ZIP4 and MER3 are required for the regulation of COs by ZEP1
- MER3~RCK, ZIP4~SPO22, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, Here, we identify the rice ZIP4 homolog, a member of the ZMM gene group, and explore its relationship with two other characterized ZMM genes, MER3 and ZEP1
- MER3~RCK, ZIP4~SPO22, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, Immunocytological analyses of wild-type rice reveal that ZIP4 presents as punctuate foci and colocalizes with MER3 in prophase I meiocytes
- MER3~RCK, ZIP4~SPO22, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, Additionally, ZIP4 is essential for the loading of MER3 onto chromosomes, but not vice versa
- MER3~RCK, ZIP4~SPO22, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, Double-mutant analyses show that zip4 mer3 displays a greater decrease in the mean number of chiasmata than either of the zip4 or mer3 single mutants, suggesting that ZIP4 and MER3 work cooperatively to promote CO formation but their individual contributions are not completely identical in rice
- MER3~RCK, ZIP4~SPO22, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, Although zep1 alone gives an increased chiasma number, both zip4 zep1 and mer3 zep1 show a much lower chiasma number than the zip4 or mer3 single mutants
- MER3~RCK, ZIP4~SPO22, ZIP4 in homologous chromosome synapsis and crossover formation in rice meiosis, These results imply that the normal functions of ZIP4 and MER3 are required for the regulation of COs by ZEP1
- MER3~RCK, OsRad21-4~OsREC8, OsSGO1 maintains synaptonemal complex stabilization in addition to protecting centromeric cohesion during rice meiosis, Finally, we found that the centromeric localization of OsSGO1 depends on OsAM1, not other meiotic proteins such as OsREC8, PAIR2, OsMER3, or ZEP1
- MER3~RCK, OsAM1, OsSGO1 maintains synaptonemal complex stabilization in addition to protecting centromeric cohesion during rice meiosis, Finally, we found that the centromeric localization of OsSGO1 depends on OsAM1, not other meiotic proteins such as OsREC8, PAIR2, OsMER3, or ZEP1
- MER3~RCK, OsSGO1, OsSGO1 maintains synaptonemal complex stabilization in addition to protecting centromeric cohesion during rice meiosis, Finally, we found that the centromeric localization of OsSGO1 depends on OsAM1, not other meiotic proteins such as OsREC8, PAIR2, OsMER3, or ZEP1
- MER3~RCK, ZEP1, OsSGO1 maintains synaptonemal complex stabilization in addition to protecting centromeric cohesion during rice meiosis, Finally, we found that the centromeric localization of OsSGO1 depends on OsAM1, not other meiotic proteins such as OsREC8, PAIR2, OsMER3, or ZEP1
- MER3~RCK, PAIR2, OsSGO1 maintains synaptonemal complex stabilization in addition to protecting centromeric cohesion during rice meiosis, Finally, we found that the centromeric localization of OsSGO1 depends on OsAM1, not other meiotic proteins such as OsREC8, PAIR2, OsMER3, or ZEP1
- MER3~RCK, OsCOM1, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Several key meiotic proteins, including ZEP1 and OsMER3, were not loaded normally onto chromosomes in Oscom1 mutants, whereas the localization of OsREC8, PAIR2 and PAIR3 seemed to be normal
- MER3~RCK, OsCOM1, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Moreover, OsCOM1 was loaded normally onto meiotic chromosomes in Osrec8, zep1 and Osmer3 mutants, but could not be properly loaded in Osam1, pair2 and OsSPO11-1(RNAi) plants
- MER3~RCK, OsSPO11-1~OsTOP6A1, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Moreover, OsCOM1 was loaded normally onto meiotic chromosomes in Osrec8, zep1 and Osmer3 mutants, but could not be properly loaded in Osam1, pair2 and OsSPO11-1(RNAi) plants
- MER3~RCK, PAIR3, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Several key meiotic proteins, including ZEP1 and OsMER3, were not loaded normally onto chromosomes in Oscom1 mutants, whereas the localization of OsREC8, PAIR2 and PAIR3 seemed to be normal
- MER3~RCK, OsRad21-4~OsREC8, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Several key meiotic proteins, including ZEP1 and OsMER3, were not loaded normally onto chromosomes in Oscom1 mutants, whereas the localization of OsREC8, PAIR2 and PAIR3 seemed to be normal
- MER3~RCK, OsRad21-4~OsREC8, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Moreover, OsCOM1 was loaded normally onto meiotic chromosomes in Osrec8, zep1 and Osmer3 mutants, but could not be properly loaded in Osam1, pair2 and OsSPO11-1(RNAi) plants
- MER3~RCK, PAIR2, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Several key meiotic proteins, including ZEP1 and OsMER3, were not loaded normally onto chromosomes in Oscom1 mutants, whereas the localization of OsREC8, PAIR2 and PAIR3 seemed to be normal
- MER3~RCK, PAIR2, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Moreover, OsCOM1 was loaded normally onto meiotic chromosomes in Osrec8, zep1 and Osmer3 mutants, but could not be properly loaded in Osam1, pair2 and OsSPO11-1(RNAi) plants
- MER3~RCK, ZEP1, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Several key meiotic proteins, including ZEP1 and OsMER3, were not loaded normally onto chromosomes in Oscom1 mutants, whereas the localization of OsREC8, PAIR2 and PAIR3 seemed to be normal
- MER3~RCK, ZEP1, The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis, Moreover, OsCOM1 was loaded normally onto meiotic chromosomes in Osrec8, zep1 and Osmer3 mutants, but could not be properly loaded in Osam1, pair2 and OsSPO11-1(RNAi) plants
- MER3~RCK, OsMSH5, The role of OsMSH5 in crossover formation during rice meiosis, OsMSH5 can be loaded onto meiotic chromosomes in Oszip4, Osmer3, and hei10
- MER3~RCK, OsMSH5, The role of OsMSH5 in crossover formation during rice meiosis, Furthermore, the residual chiasmata were shown to be the least frequent among the zmm mutants, including Osmer3, Oszip4, hei10, and Osmsh5
- MER3~RCK, OsMSH5, The role of OsMSH5 in crossover formation during rice meiosis, Taken together, we propose that OsMSH5 functions upstream of OsZIP4, OsMER3, and HEI10 in class I crossover formation
- HEI10, MER3~RCK, The role of OsMSH5 in crossover formation during rice meiosis, OsMSH5 can be loaded onto meiotic chromosomes in Oszip4, Osmer3, and hei10
- HEI10, MER3~RCK, The role of OsMSH5 in crossover formation during rice meiosis, Furthermore, the residual chiasmata were shown to be the least frequent among the zmm mutants, including Osmer3, Oszip4, hei10, and Osmsh5
- HEI10, MER3~RCK, The role of OsMSH5 in crossover formation during rice meiosis, Taken together, we propose that OsMSH5 functions upstream of OsZIP4, OsMER3, and HEI10 in class I crossover formation
- MER3~RCK, ZIP4~SPO22, The role of OsMSH5 in crossover formation during rice meiosis, OsMSH5 can be loaded onto meiotic chromosomes in Oszip4, Osmer3, and hei10
- MER3~RCK, ZIP4~SPO22, The role of OsMSH5 in crossover formation during rice meiosis, Furthermore, the residual chiasmata were shown to be the least frequent among the zmm mutants, including Osmer3, Oszip4, hei10, and Osmsh5
- MER3~RCK, ZIP4~SPO22, The role of OsMSH5 in crossover formation during rice meiosis, Taken together, we propose that OsMSH5 functions upstream of OsZIP4, OsMER3, and HEI10 in class I crossover formation
- MER3~RCK, PAIR2, The central element protein ZEP1 of the synaptonemal complex regulates the number of crossovers during meiosis in rice, Although PAIR2 and MER3 were loaded normally in zep1, their dissociation was delayed severely compared with the wild type
- MER3~RCK, ZEP1, The central element protein ZEP1 of the synaptonemal complex regulates the number of crossovers during meiosis in rice, Although PAIR2 and MER3 were loaded normally in zep1, their dissociation was delayed severely compared with the wild type
- MER3~RCK, OsRad21-4~OsREC8, OsREC8 is essential for chromatid cohesion and metaphase I monopolar orientation in rice meiosis, Immunolocalization analyses revealed that the loading of PAIR2, PAIR3, OsMER3, and ZEP1 all depended on OsREC8
- MER3~RCK, OsRad21-4~OsREC8, OsREC8 is essential for chromatid cohesion and metaphase I monopolar orientation in rice meiosis, By contrast, the presence of the OsREC8 signal in pair2, pair3, Osmer3, and zep1 mutants indicated that the loading of OsREC8 did not rely on these four proteins
- MER3~RCK, PAIR2, OsREC8 is essential for chromatid cohesion and metaphase I monopolar orientation in rice meiosis, Immunolocalization analyses revealed that the loading of PAIR2, PAIR3, OsMER3, and ZEP1 all depended on OsREC8
- MER3~RCK, PAIR2, OsREC8 is essential for chromatid cohesion and metaphase I monopolar orientation in rice meiosis, By contrast, the presence of the OsREC8 signal in pair2, pair3, Osmer3, and zep1 mutants indicated that the loading of OsREC8 did not rely on these four proteins
- MER3~RCK, ZEP1, OsREC8 is essential for chromatid cohesion and metaphase I monopolar orientation in rice meiosis, Immunolocalization analyses revealed that the loading of PAIR2, PAIR3, OsMER3, and ZEP1 all depended on OsREC8
- MER3~RCK, ZEP1, OsREC8 is essential for chromatid cohesion and metaphase I monopolar orientation in rice meiosis, By contrast, the presence of the OsREC8 signal in pair2, pair3, Osmer3, and zep1 mutants indicated that the loading of OsREC8 did not rely on these four proteins
- MER3~RCK, XRCC3, XRCC3 is essential for proper double-strand break repair and homologous recombination in rice meiosis., Moreover, abnormal chromosome localization of RAD51C, DMC1, ZEP1, ZIP4, and MER3 was observed in xrcc3
Prev Next